Flavanol Polymerization Is a Superior Predictor of α-Glucosidase Inhibitory Activity Compared to Flavanol or Total Polyphenol Concentrations in Cocoas Prepared by Variations in Controlled Fermentation and Roasting of the Same Raw Cocoa Beans

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Standards
2.2. Fermentation Model System and Processing
2.2.1. Fermentation
2.2.2. Roasting and Further Processing
2.3. Fermentation Assays
2.4. Polyphenol Extraction and Characterization
2.5. Melanoidin Dialysis
2.6. In Vitro α-Glucosidase Inhibition Assay
- ΔAsample= the change in the individual absorbance value of the product of the inhibitor, substrate, and enzyme at each inhibitor dose before and after incubation; and,
- Δnegative control= the average change in absorbance of the negative control (0 µg/mL) before and after incubation.
2.7. Data Analysis and Statistics
3. Results
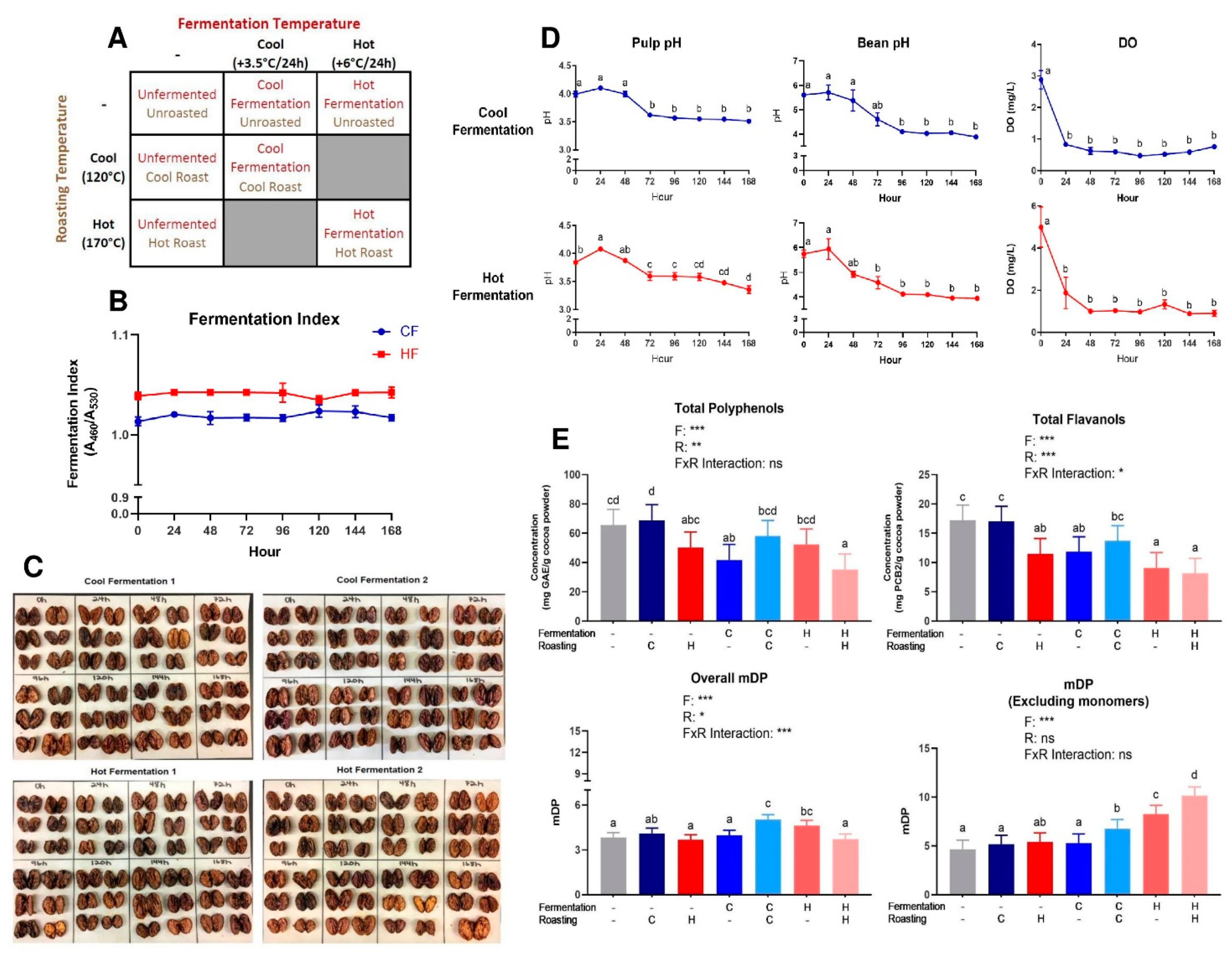
3.1. Fermentation Model System and Cocoa Processing
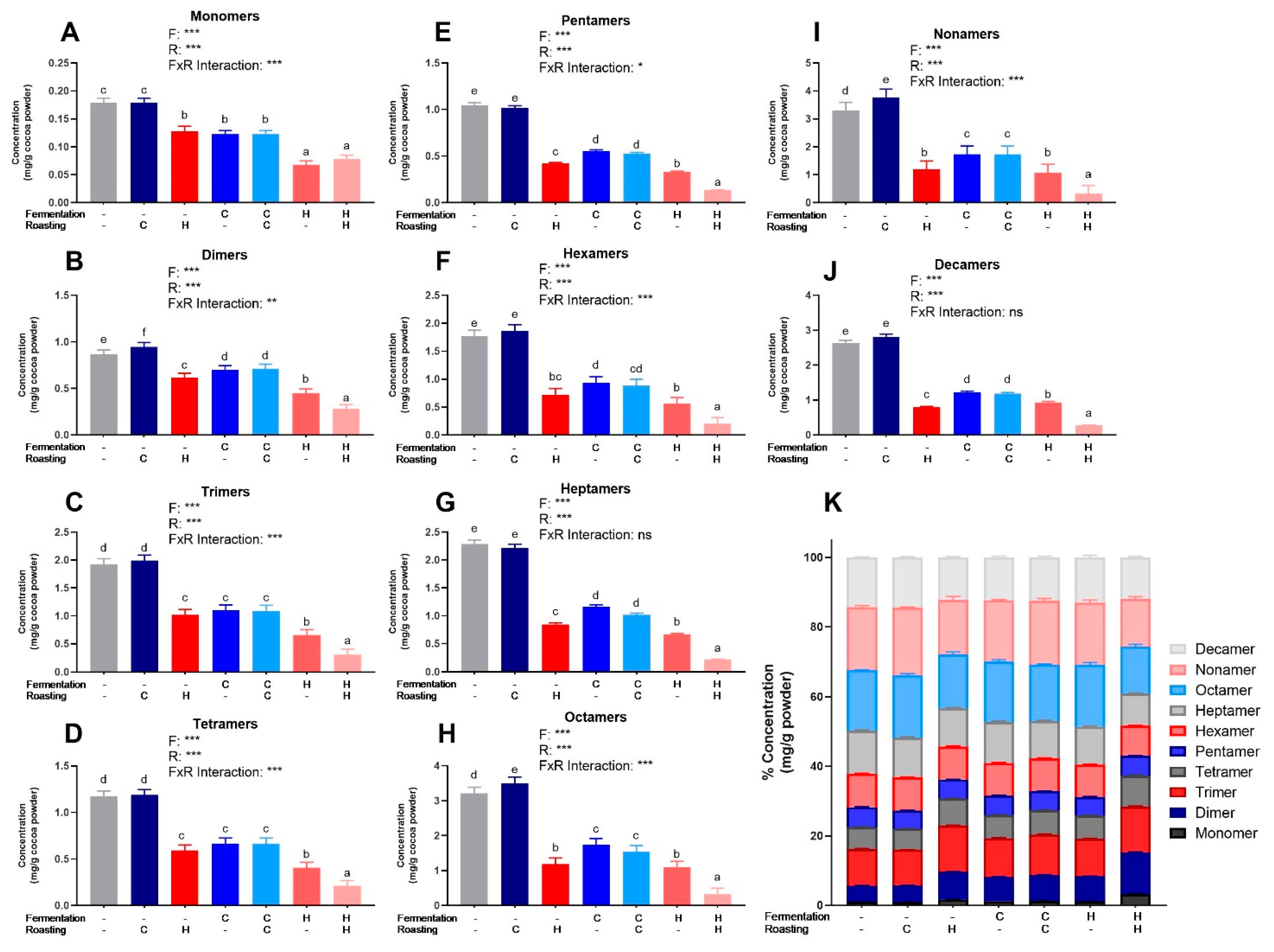
3.2. Characterization and Quantification of Polyphenols
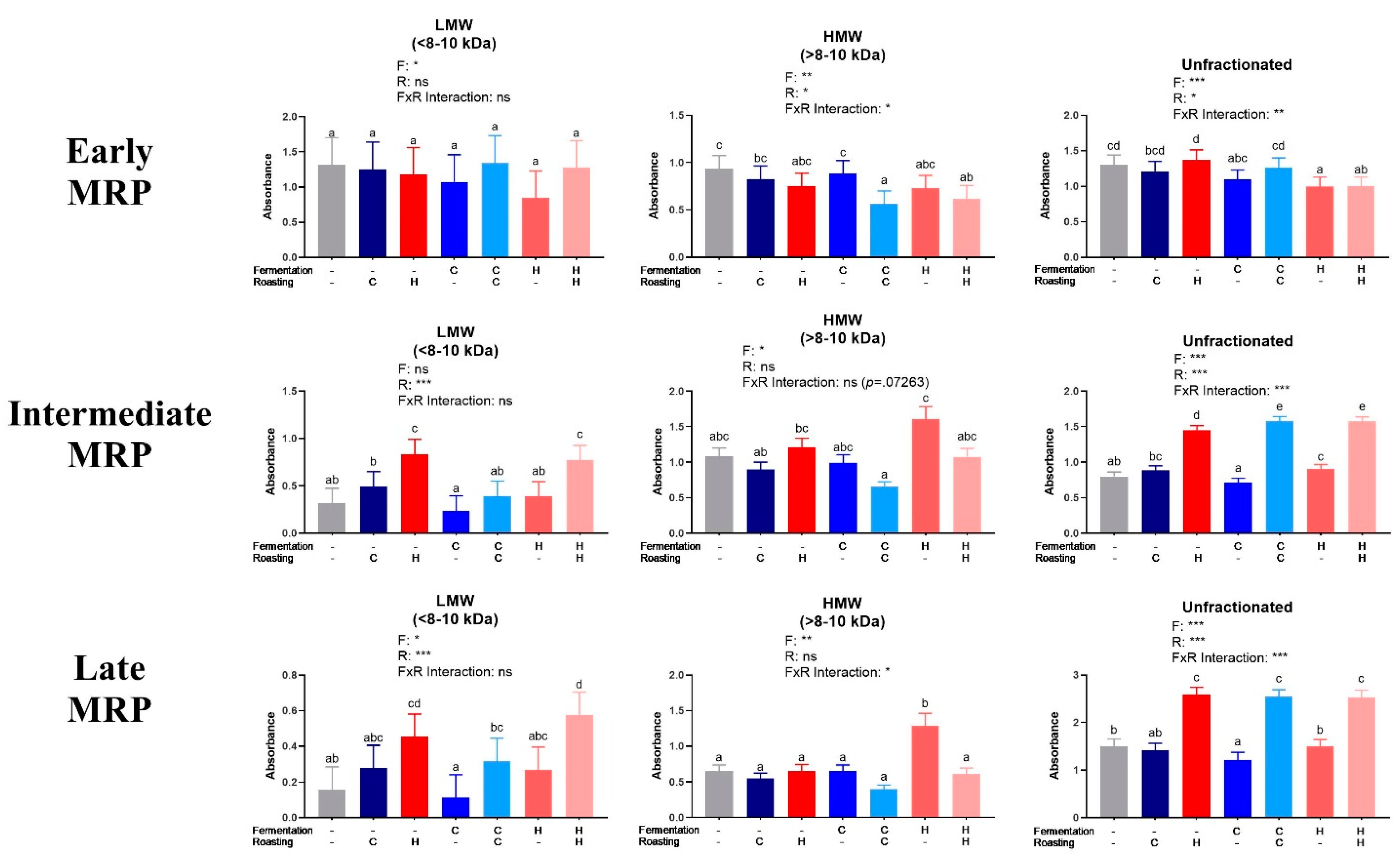
3.3. Quantification of Maillard Reaction Products
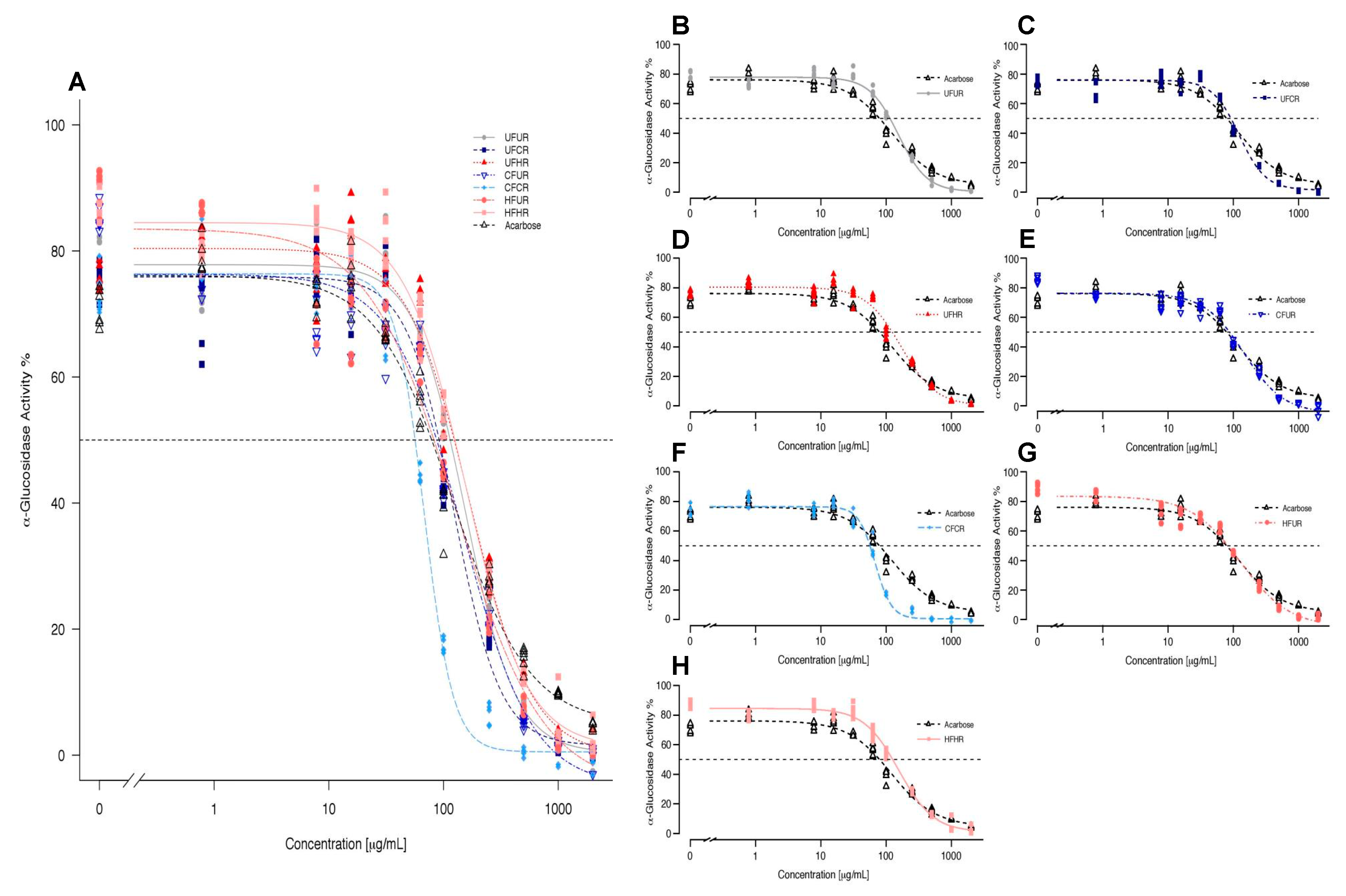
3.4. α-Glucosidase Enzyme Inhibition
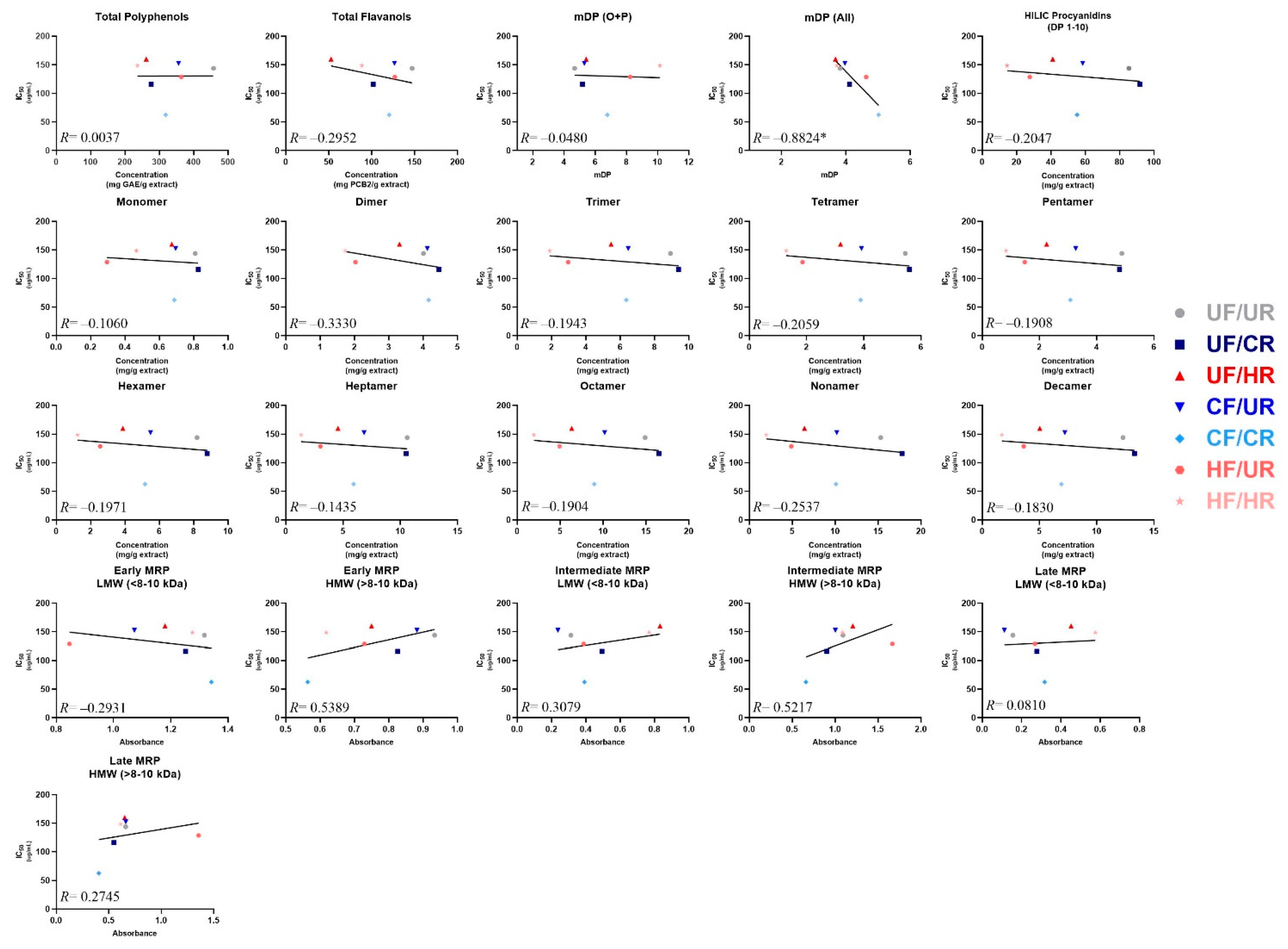
3.5. Identifying Predictors of α-Glucosidase Enzyme Inhibitory Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andújar, I.; Recio, M.C.; Giner, R.M.; Ríos, J.L. Cocoa polyphenols and their potential benefits for human health. Oxid. Med. Cell. Longev. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Kongor, J.E.; Hinneh, M.; de Walle, D.V.; Afoakwa, E.O.; Boeckx, P.; Dewettinck, K. Factors influencing quality variation in cocoa (Theobroma cacao) bean flavour profile—A review. Food Res. Int. 2016, 82, 44–52. [Google Scholar] [CrossRef]
- Schinella, G.; Mosca, S.; Cienfuegos-Jovellanos, E.; Pasamar, M.A.; Muguerza, B.; Ramon, D.; Rios, J.L. Antioxidant properties of polyphenol-rich cocoa products industrially processed. Food Res. Int. 2010, 43, 1614–1623. [Google Scholar] [CrossRef] [Green Version]
- Hatano, T.; Miyatake, H.; Natsume, M.; Osakabe, N.; Takizawa, T.; Ito, H.; Yoshida, T. Proanthocyanidin glycosides and related polyphenols from cacao liquor and their antioxidant effects. Phytochemistry 2002, 59, 749–758. [Google Scholar] [CrossRef]
- Oracz, J.; Nebesny, E.; Żyżelewicz, D. Changes in the flavan-3-ols, anthocyanins, and flavanols composition of cocoa beans of different Theobroma cacao L. groups affected by roasting conditions. Eur. Food Res. Technol. 2015, 241, 663–681. [Google Scholar] [CrossRef] [Green Version]
- Cooper, K.A.; Donovan, J.L.; Waterhouse, A.L.; Williamson, G. Cocoa and health: A decade of research. Br. J. Nutr. 2008, 99, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wollgast, J.; Anklam, E. Review on polyphenols in Theobroma cacao: Changes in composition during the manufacture of chocolate and methodology for identification and quantification. Food Res. Int. 2000, 33, 423–447. [Google Scholar] [CrossRef]
- de Brito, E.S.; García, N.H.P.; Gallão, M.I.; Cortelazzo, A.L.; Fevereiro, P.S.; Braga, M.R. Structural and chemical changes in cocoa (Theobroma cacao L) during fermentation, drying and roasting. J. Sci. Food Agric. 2001, 81, 281–288. [Google Scholar] [CrossRef]
- Mazor Jolić, S.; Radojčić Redovniković, I.; Marković, K.; Ivanec Šipušić, Đ.; Delonga, K. Changes of phenolic compounds and antioxidant capacity in cocoa beans processing. Int. J. Food Sci. Technol. 2011, 46, 1793–1800. [Google Scholar] [CrossRef]
- Oracz, J.; Nebesny, E. Effect of roasting parameters on the physicochemical characteristics of high-molecular-weight Maillard reaction products isolated from cocoa beans of different Theobroma cacao L. groups. Eur. Food Res. Technol. 2019, 245, 111–128. [Google Scholar] [CrossRef]
- Albertini, B.; Schoubben, A.; Guarnaccia, D.; Pinelli, F.; Della Vecchia, M.; Ricci, M.; Di Renzo, G.C.; Blasi, P. Effect of fermentation and drying on cocoa polyphenols. J. Agric. Food Chem. 2015, 63, 9948–9953. [Google Scholar] [CrossRef] [PubMed]
- Dorenkott, M.R.; Griffin, L.E.; Goodrich, K.M.; Thompson-Witrick, K.A.; Fundaro, G.; Ye, L.; Stevens, J.R.; Ali, M.; O’Keefe, S.F.; Hulver, M.W.; et al. Oligomeric Cocoa Procyanidins Possess Enhanced Bioactivity Compared to Monomeric and Polymeric Cocoa Procyanidins for Preventing the Development of Obesity, Insulin Resistance, and Impaired Glucose Tolerance during High-Fat Feeding. J. Agric. Food Chem. 2014, 62, 2216–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, C.M.; Khoo, W.; Ye, L.; Lambert, J.D.; O’Keefe, S.F.; Neilson, A.P. Loss of Native Flavanols during Fermentation and Roasting Does Not Necessarily Reduce Digestive Enzyme-Inhibiting Bioactivities of Cocoa. J. Agric. Food Chem. 2016, 64, 3616–3625. [Google Scholar] [CrossRef] [PubMed]
- Di Mattia, C.; Martuscelli, M.; Sacchetti, G.; Scheirlinck, I.; Beheydt, B.; Mastrocola, D.; Pittia, P. Effect of Fermentation and Drying on Procyanidins, Antiradical Activity and Reducing Properties of Cocoa Beans. Food Bioprocess Technol. 2013, 6, 3420–3432. [Google Scholar] [CrossRef]
- Quiroz-Reyes, C.N.; Fogliano, V. Design cocoa processing towards healthy cocoa products: The role of phenolics and melanoidins. J. Funct. Foods 2018, 45, 480–490. [Google Scholar] [CrossRef]
- Bellesia, A.; Tagliazucchi, D. Cocoa brew inhibits in vitro α-glucosidase activity: The role of polyphenols and high molecular weight compounds. Food Res. Int. 2014, 63, 439–445. [Google Scholar] [CrossRef]
- Ioannone, F.; Di Mattia, C.D.; De Gregorio, M.; Sergi, M.; Serafini, M.; Sacchetti, G. Flavanols, proanthocyanidins and antioxidant activity changes during cocoa (Theobroma cacao L.) roasting as affected by temperature and time of processing. Food Chem. 2015, 174, 256–262. [Google Scholar] [CrossRef]
- Summa, C.; McCourt, J.; Cämmerer, B.; Fiala, A.; Probst, M.; Kun, S.; Anklam, E.; Wagner, K.-H. Radical scavenging activity, anti-bacterial and mutagenic effects of Cocoa bean Maillard Reaction products with degree of roasting. Mol. Nutr. Food Res. 2008, 52, 342–351. [Google Scholar] [CrossRef]
- Summa, C.; Raposo, F.C.; McCourt, J.; Scalzo, R.L.; Wagner, K.-H.; Elmadfa, I.; Anklam, E. Effect of roasting on the radical scavenging activity of cocoa beans. Eur. Food Res. Technol. 2006, 222, 368–375. [Google Scholar] [CrossRef]
- Donovan, J.L.; Lee, A.; Manach, C.; Rios, L.; Morand, C.; Scalbert, A.; Rémésy, C. Procyanidins are not bioavailable in rats fed a single meal containing a grapeseed extract or the procyanidin dimer B 3. Br. J. Nutr. 2002, 87, 299–306. [Google Scholar] [CrossRef]
- Serra, A.; Macia, A.; Romero, M.-P.; Valls, J.; Bladé, C.; Arola, L.; Motilva, M.-J. Bioavailability of procyanidin dimers and trimers and matrix food effects in in vitro and in vivo models. Br. J. Nutr. 2010, 103, 944–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottaviani, J.I.; Kwik-Uribe, C.; Keen, C.L.; Schroeter, H. Intake of dietary procyanidins does not contribute to the pool of circulating flavanols in humans. Am. J. Clin. Nutr. 2012, 95, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crozier, A.; Del Rio, D.; Clifford, M.N. Bioavailability of dietary flavonoids and phenolic compounds. Mol. Aspects Med. 2010, 31, 446–467. [Google Scholar] [CrossRef] [PubMed]
- Wiese, S.; Esatbeyoglu, T.; Winterhalter, P.; Kruse, H.-P.; Winkler, S.; Bub, A.; Kulling, S.E. Comparative biokinetics and metabolism of pure monomeric, dimeric, and polymeric flavan-3-ols: A randomized cross-over study in humans. Mol. Nutr. Food Res. 2015, 59, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Hurst, W.J.; Stuart, D.A.; Lambert, J.D. Inhibition of Key Digestive Enzymes by Cocoa Extracts and Procyanidins. J. Agric. Food Chem. 2011, 59, 5305–5311. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.-L.; Han, L.; Yang, S.-Y.; Meng, X.-M.; Ma, W.-F.; Wang, M. The mechanism of interactions between flavan-3-ols against a-glucosidase and their in vivo antihyperglycemic effects. Bioorganic Chem. 2019, 85, 364–372. [Google Scholar] [CrossRef]
- Camu, N.; Winter, T.D.; Verbrugghe, K.; Cleenwerck, I.; Vandamme, P.; Takrama, J.S.; Vancanneyt, M.; Vuyst, L.D. Dynamics and Biodiversity of Populations of Lactic Acid Bacteria and Acetic Acid Bacteria Involved in Spontaneous Heap Fermentation of Cocoa Beans in Ghana. Appl. Environ. Microbiol. 2007, 73, 1809–1824. [Google Scholar] [CrossRef] [Green Version]
- Camu, N.; González, A.; De Winter, T.; Van Schoor, A.; De Bruyne, K.; Vandamme, P.; Takrama, J.S.; Addo, S.K.; De Vuyst, L. Influence of turning and environmental contamination on the dynamics of populations of lactic acid and acetic acid bacteria involved in spontaneous cocoa bean heap fermentation in Ghana. Appl. Environ. Microbiol. 2008, 74, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Schwan, R.F.; Wheals, A.E. The Microbiology of Cocoa Fermentation and its Role in Chocolate Quality. Crit. Rev. Food Sci. Nutr. 2004, 44, 205–221. [Google Scholar] [CrossRef]
- Racine, K.C.; Lee, A.H.; Wiersema, B.D.; Huang, H.; Lambert, J.D.; Stewart, A.C.; Neilson, A.P. Development and Characterization of a Pilot-Scale Model Cocoa Fermentation System Suitable for Studying the Impact of Fermentation on Putative Bioactive Compounds and Bioactivity of Cocoa. Foods 2019, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.H.; Neilson, A.P.; O’Keefe, S.F.; Ogejo, J.A.; Huang, H.; Ponder, M.; Chu, H.S.S.; Jin, Q.; Pilot, G.; Stewart, A.C. A laboratory-scale model cocoa fermentation using dried, unfermented beans and artificial pulp can simulate the microbial and chemical changes of on-farm cocoa fermentation. Eur. Food Res. Technol. 2018. [Google Scholar] [CrossRef]
- Camu, N.; De Winter, T.; Addo, S.K.; Takrama, J.S.; Bernaert, H.; De Vuyst, L. Fermentation of cocoa beans: influence of microbial activities and polyphenol concentrations on the flavour of chocolate. J. Sci. Food Agric. 2008, 88, 2288–2297. [Google Scholar] [CrossRef]
- Schwan, R.F. Cocoa fermentations conducted with a defined microbial cocktail inoculum. Appl. Environ. Microbiol. 1998, 64, 1477–1483. [Google Scholar] [PubMed]
- Romero-Cortes, T.; Salgado-Cervantes, M.A.; García-Alamilla, P.; García-Alvarado, M.A.; del C Rodríguez-Jimenes, G.; Hidalgo-Morales, M.; Robles-Olvera, V. Relationship between fermentation index and other biochemical changes evaluated during the fermentation of Mexican cocoa (Theobroma cacao) beans. J. Sci. Food Agric. 2013, 93, 2596–2604. [Google Scholar] [CrossRef]
- Racine, K.C.; Lee, A.H.; Stewart, A.C.; Blakeslee, K.W.; Neilson, A.P. Development of a rapid ultra performance hydrophilic interaction liquid chromatography tandem mass spectrometry method for procyanidins with enhanced ionization efficiency. J. Chromatogr. A 2019, 1594, 54–64. [Google Scholar] [CrossRef]
- Sacchetti, G.; Ioannone, F.; De Gregorio, M.; Di Mattia, C.; Serafini, M.; Mastrocola, D. Non enzymatic browning during cocoa roasting as affected by processing time and temperature. J. Food Eng. 2016, 169, 44–52. [Google Scholar] [CrossRef]
- Balfour, J.A.; McTavish, D. Acarbose. Drugs 1993, 46, 1025–1054. [Google Scholar] [CrossRef]
- Nielsen, D.S.; Teniola, O.D.; Ban-Koffi, L.; Owusu, M.; Andersson, T.S.; Holzapfel, W.H. The microbiology of Ghanaian cocoa fermentations analysed using culture-dependent and culture-independent methods. Int. J. Food Microbiol. 2007, 114, 168–186. [Google Scholar] [CrossRef]
- Ho, V.T.T.; Zhao, J.; Fleet, G. The effect of lactic acid bacteria on cocoa bean fermentation. Int. J. Food Microbiol. 2015, 205, 54–67. [Google Scholar] [CrossRef]
- Ardhana, M.M.; Fleet, G.H. The microbial ecology of cocoa bean fermentations in Indonesia. Int. J. Food Microbiol. 2003, 86, 87–99. [Google Scholar] [CrossRef]
- Saltini, R.; Akkerman, R.; Frosch, S. Optimizing chocolate production through traceability: A review of the influence of farming practices on cocoa bean quality. Food Control 2013, 29, 167–187. [Google Scholar] [CrossRef] [Green Version]
- De Taeye, C.; Eyamo Evina, V.J.; Caullet, G.; Niemenak, N.; Collin, S. Fate of anthocyanins through cocoa fermentation. Emergence of new polyphenolic dimers. J. Agric. Food Chem. 2016, 64, 8876–8885. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, O.A.; Jennifer, Q.; Agnes, S.B.; Jemmy, S.T.; Firibu, K.S. Influence of pulp-preconditioning and fermentation on fermentative quality and appearance of Ghanaian cocoa (Theobroma cacao) beans. Int. Food Res. J. Selangor. 2012, 19, 127–133. [Google Scholar]
- Payne, M.J.; Hurst, W.J.; Miller, K.B.; Rank, C.; Stuart, D.A. Impact of Fermentation, Drying, Roasting, and Dutch Processing on Epicatechin and Catechin Content of Cacao Beans and Cocoa Ingredients. J. Agric. Food Chem. 2010, 58, 10518–10527. [Google Scholar] [CrossRef]
- Kim, H.; Keeney, P.G. (−)-Epicatechin Content in Fermented and Unfermented Cocoa Beans. J. Food Sci. 1984, 49, 1090–1092. [Google Scholar] [CrossRef]
- Kothe, L.; Zimmermann, B.F.; Galensa, R. Temperature influences epimerization and composition of flavanol monomers, dimers and trimers during cocoa bean roasting. Food Chem. 2013, 141, 3656–3663. [Google Scholar] [CrossRef]
- Ryan, C.M.; Khoo, W.; Stewart, A.C.; O’Keefe, S.F.; Lambert, J.D.; Neilson, A.P. Flavanol concentrations do not predict dipeptidyl peptidase-IV inhibitory activities of four cocoas with different processing histories. Food Funct. 2017, 8, 746–756. [Google Scholar] [CrossRef]
- Zhong, H.; Xue, Y.; Lu, X.; Shao, Q.; Cao, Y.; Wu, Z.; Chen, G. The Effects of Different Degrees of Procyanidin Polymerization on the Nutrient Absorption and Digestive Enzyme Activity in Mice. Molecules 2018, 23, 2916. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, Y.; Okabe, M.; Natsume, M.; Ashida, H. Comparison of anti-hyperglycemic activities between low-and high-degree of polymerization procyanidin fractions from cacao liquor extract. J. Food Drug Anal. 2012, 20, 283–287. [Google Scholar]
- Hogan, S.; Zhang, L.; Li, J.; Sun, S.; Canning, C.; Zhou, K. Antioxidant rich grape pomace extract suppresses postprandial hyperglycemia in diabetic mice by specifically inhibiting alpha-glucosidase. Nutr. Metab. 2010, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Lavelli, V.; Harsha, P.S.; Ferranti, P.; Scarafoni, A.; Iametti, S. Grape skin phenolics as inhibitors of mammalian α-glucosidase and α-amylase–effect of food matrix and processing on efficacy. Food Funct. 2016, 7, 1655–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougall, G.J.; Shpiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different polyphenolic components of soft fruits inhibit α-amylase and α-glucosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent e | Mass a (g) |
|---|---|
| Simulated Pulp Solution b | |
| Citric acid | 40 |
| Yeast extract | 20 |
| Peptone | 20 |
| Calcium lactate pentahydrate | 4 |
| Tween 80 | 4 |
| Magnesium-Manganese Solution c | |
| Magnesium sulfate-heptahydride | 2 |
| Manganese sulfate-monohydride | 0.8 |
| Sugar Solutions d | |
| Sucrose | 100 |
| Glucose | 160 |
| Fructose | 180 |
| Parameter | Treatment | Estimate | Std. Error | LCL (2.5%) | UCL (97.5%) | t-Value | p-Value | Significance | ||
|---|---|---|---|---|---|---|---|---|---|---|
| F | R | Abbreviation | ||||||||
| Hill Coefficient | - | - | (UF/UR) | 1.9696 | 0.141725 | 1.691 | 2.248 | 13.8973 | <2.2 × 10−16 | *** |
| - | Cool | (UF/CR) | 2.142171 | 0.177728 | 1.793 | 2.491 | 12.0531 | <2.2 × 10−16 | *** | |
| - | Hot | (UF/HR) | 1.603793 | 0.113242 | 1.381 | 1.826 | 14.1626 | <2.2 × 10−16 | *** | |
| Cool | - | (CF/UR) | 1.429009 | 0.129739 | 1.174 | 1.684 | 11.0145 | <2.2 × 10−16 | *** | |
| Cool | Cool | (CF/CR) | 3.465039 | 0.425681 | 2.629 | 4.301 | 8.14 | 3.17 × 10−16 | *** | |
| Hot | - | (HF/UR) | 1.142063 | 0.107241 | 0.931 | 1.353 | 10.6495 | <2.2 × 10−16 | *** | |
| Hot | Hot | (HF/HR) | 1.627053 | 0.113836 | 1.403 | 1.851 | 14.2929 | <2.2 × 10−16 | *** | |
| Acarbose | (A) | 1.30504 | 0.113057 | 1.083 | 1.527 | 11.5432 | <2.2 × 10−16 | *** | ||
| Minimum Value | - | - | (UF/UR) | 0.378196 | 1.496104 | −2.561 | 3.318 | 0.2528 | 0.80054 | |
| - | Cool | (UF/CR) | 1.513805 | 1.388439 | −1.214 | 4.242 | 1.0903 | 0.27611 | ||
| - | Hot | (UF/HR) | −0.030323 | 1.857523 | −3.68 | 3.619 | −0.0163 | 0.98698 | ||
| Cool | - | (CF/UR) | −4.888001 | 2.078745 | −8.972 | −0.804 | −2.3514 | 0.01909 | * | |
| Cool | Cool | (CF/CR) | 0.516432 | 0.977596 | −1.404 | 2.437 | 0.5283 | 0.59755 | ||
| Hot | - | (HF/UR) | −5.208691 | 2.488998 | −10.1 | −0.318 | −2.0927 | 0.03689 | * | |
| Hot | Hot | (HF/HR) | 1.208122 | 1.755526 | −2.241 | 4.657 | 0.6882 | 0.49166 | ||
| Acarbose | (A) | 4.867636 | 2.043044 | 0.854 | 8.882 | 2.3825 | 0.01757 | * | ||
| Maximum Value | - | - | (UF/UR) | 77.81598 | 0.894435 | 76.059 | 79.573 | 87.0002 | <2.2 × 10−16 | *** |
| - | Cool | (UF/CR) | 75.880224 | 0.889772 | 74.132 | 77.628 | 85.2806 | <2.2 × 10−16 | *** | |
| - | Hot | (UF/HR) | 80.391462 | 0.940686 | 78.543 | 82.24 | 85.4604 | <2.2 × 10−16 | *** | |
| Cool | - | (CF/UR) | 76.297669 | 1.107626 | 74.121 | 78.474 | 68.8839 | <2.2 × 10−16 | *** | |
| Cool | Cool | (CF/CR) | 76.360965 | 0.968329 | 74.458 | 78.264 | 78.8585 | <2.2 × 10−16 | *** | |
| Hot | - | (HF/UR) | 83.518081 | 1.382998 | 80.801 | 86.235 | 60.3892 | <2.2 × 10−16 | *** | |
| Hot | Hot | (HF/HR) | 84.494321 | 0.96061 | 82.607 | 86.382 | 87.959 | <2.2 × 10−16 | *** | |
| Acarbose | (A) | 76.123584 | 1.073488 | 74.014 | 78.233 | 70.9123 | <2.2 × 10−16 | *** | ||
| EC50 | - | - | (UF/UR) | 153.38225 | 8.067667 | 137.53 | 169.23 | 19.012 | <2.2 × 10−16 | *** |
| - | Cool | (UF/CR) | 122.95011 | 6.486545 | 110.21 | 135.7 | 18.9546 | <2.2 × 10−16 | *** | |
| - | Hot | (UF/HR) | 169.26537 | 10.618809 | 148.4 | 190.13 | 15.9401 | <2.2 × 10−16 | *** | |
| Cool | - | (CF/UR) | 146.08776 | 9.519332 | 127.38 | 164.79 | 15.3464 | <2.2 × 10−16 | *** | |
| Cool | Cool | (CF/CR) | 67.824539 | 1.888888 | 64.113 | 71.536 | 35.9071 | <2.2 × 10−16 | *** | |
| Hot | - | (HF/UR) | 127.62786 | 9.183673 | 109.58 | 145.67 | 13.8973 | <2.2 × 10−16 | *** | |
| Hot | Hot | (HF/HR) | 155.5339 | 8.935954 | 137.98 | 173.09 | 17.4054 | <2.2 × 10−16 | *** | |
| Acarbose | (A) | 119.95973 | 10.225587 | 99.869 | 140.05 | 11.7313 | <2.2 × 10−16 | *** | ||
| IC50 a | - | - | (UF/UR) | 154.1448 | - | 132.33 a | 175.92 b | - | - | - |
| - | Cool | (UF/CR) | 125.30945 | - | 108.24 a | 142.15 b | - | - | - | |
| - | Hot | (UF/HR) | 169.1858 | - | 139.08 a | 199.97 b | - | - | - | |
| Cool | - | (CF/UR) | 134.26837 | - | 105.91 a | 162.82 b | - | - | - | |
| Cool | Cool | (CF/CR) | 68.09163 | - | 63.22 a | 72.61 b | - | - | - | |
| Hot | - | (HF/UR) | 115.14538 | - | 86.2 4 a | 144.88 b | - | - | - | |
| Hot | Hot | (HF/HR) | 158.33224 | - | 132.88 a | 184.1 b | - | - | - | |
| Acarbose | (A) | 133.22093 | - | 102.04 a | 165.78 b | - | - | - | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Racine, K.C.; Wiersema, B.D.; Griffin, L.E.; Essenmacher, L.A.; Lee, A.H.; Hopfer, H.; Lambert, J.D.; Stewart, A.C.; Neilson, A.P. Flavanol Polymerization Is a Superior Predictor of α-Glucosidase Inhibitory Activity Compared to Flavanol or Total Polyphenol Concentrations in Cocoas Prepared by Variations in Controlled Fermentation and Roasting of the Same Raw Cocoa Beans. Antioxidants 2019, 8, 635. https://doi.org/10.3390/antiox8120635
Racine KC, Wiersema BD, Griffin LE, Essenmacher LA, Lee AH, Hopfer H, Lambert JD, Stewart AC, Neilson AP. Flavanol Polymerization Is a Superior Predictor of α-Glucosidase Inhibitory Activity Compared to Flavanol or Total Polyphenol Concentrations in Cocoas Prepared by Variations in Controlled Fermentation and Roasting of the Same Raw Cocoa Beans. Antioxidants. 2019; 8(12):635. https://doi.org/10.3390/antiox8120635
Chicago/Turabian StyleRacine, Kathryn C., Brian D. Wiersema, Laura E. Griffin, Lauren A. Essenmacher, Andrew H. Lee, Helene Hopfer, Joshua D. Lambert, Amanda C. Stewart, and Andrew P. Neilson. 2019. "Flavanol Polymerization Is a Superior Predictor of α-Glucosidase Inhibitory Activity Compared to Flavanol or Total Polyphenol Concentrations in Cocoas Prepared by Variations in Controlled Fermentation and Roasting of the Same Raw Cocoa Beans" Antioxidants 8, no. 12: 635. https://doi.org/10.3390/antiox8120635